Есть ли днк у бактерий. Генетический материал бактерий. Днк бактерии, днк бактерия в мазке
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.
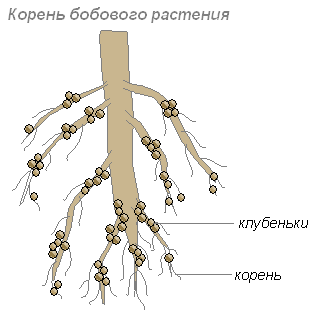
Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
ГЕНЕТИКА БАКТЕРИЙ
Цель лекции: ознакомить студентов с основами генетики бактерий, с основными методами генодиагностики.
План лекции
1. Особенности организации ядерного аппарата бактерий.
2. Состав бактериального генома.
3. Изменчивость бактерий.
4. Рекомбинации у бактерий и их особенности.
5. Генодиагностика.
6. Классификация бактерий.
1.Особенности морфологической организации ядерного аппарата бактерий:
- не имеет ядерной мембраны, ядрышка; носит название нуклеоид;
- носителем генетической информации является ДНК. Если у эукариот ДНКлинейная, то у большинства бактерий - кольцевая, и одна нить фиксирована на цитоплазматической мембране. Если раскрутить ДНК, то длина её будет в сотни раз превышать длину клетки. ДНК бактерий суперспирализована.
- бактериальная клетка содержит одну хромосому, т.е. бактерии являются гаплоидными организмами.
2. Биохимические особенности .
- ДНК бактерий имеет тот же состав, что и ДНК эукариот.
- у бактерий в составе ДНК могут находиться минорные основания, наличие которых защищает ДНК от действия собственных эндонуклеаз.
- в геноме патогенных бактерий имеются участки ДНК, которые отличаются от основного генома составом Г-Ц пар нуклеотидных оснований. Эти участки ответственны за синтез факторов патогенности-острова патогенности.
- ДНК бактерий не содержит гистонов, а их роль выполняют полиамины. Бактериальный геном представлен структурами, которые способны к
автономной репликации. Таких структур две: хромосомы , в которых закодирована вся жизненно необходимая информация (в хромосоме бактерий содержится до 3 тыс. различных генов), и плазмиды .
Плазмиды - это ДНК, которые имеют кольцевую природу. Плазмиды в клетке могут находиться в одном из двух альтернативных состояний: в свободном или интегрированном с хромосомой.
В плазмидах закодирована дополнительная генетическая информация, которая не является жизненно необходимой для клетки, но наличие этой информации сообщает ей определенные селективные преимущества. В состав плазмид входят:
Структурные гены; -гены, отвечающие за собственную репликацию плазмиды.
Некоторые плазмиды имеют гены, обеспечивающие трансмиссивность плазмиды - tra-гены.
По кодируемому признаку различают:
- R плазмидыкодируют лекарственную устойчивость бактерий;
- F (sex) плазмиды - определяют способность клетки быть донором генетической информации;
- Col плазмида - кодирует синтез бактериоцинов;
- плазмиды, отвечающие за синтез факторов вирулентности (Ent-, Hly-)
и другие плазмиды.
В состав бактериального генома входят подвижные генетические элементы: IS-элементы (insertion sequences), транспозоны и интегроны. Они обнаружены как в составе бактериальной хромосомы, так и в составе плазмид. Их репликация – составная часть репликации хромосомы и плазмиды.
IS-элементы - короткие (2000) нуклеотидные последовательности. Отличительной особенностью IS-элементов является наличие на концах инвертированных повторов, которые узнает транспозаза. Они не несут структурных генов; одинаковы у бактерий разных видов, родов, и даже считается, что они одинаковы у прокариот и у эукариот. IS-элементы могут перемещаться как по хромосоме, так и между хромосомами. Они содержат 2 гена:1-й кодирует синтез транспозазы; этот фермент обеспечивает процесс исключения IS элемента из хромосомы и его интеграцию в новой локус хромосомы. 2-й ген кодирует синтез репрессора, который регулирует весь процесс перемещения.
Транспозоны – это сегменты ДНК, обладающие теми же свойствами, что и ISэлемент, но имеющие структурные гены.
Интегроны – подвижные генетические элементы; они содержат ген, кодирующий устойчивость к антибиотикам. Интегроны являются системой захвата малых элементов ДНК, называемых генными кассетами посредством сайтспецифической рекомбинации и их экспрессии.
Значение мобильных элементов.
Перемещаясь по ДНК клетки или между ДНК разных клеток, они вызывают:
- инактивацию генов тех участков ДНК, куда они, переместившись, встраиваются;
- повреждения генетического материала;
- встраивание плазмиды в хромосому;
- распространение гена в популяции бактерий.
Бактериям, как и всем живым существам, свойственна изменчивость. Изменчивость у эукариот происходит по вертикали, у бактерий – и по вертикали, и по горизонтали.
Различают два вида изменчивости: - фенотипическая -генотипическая.
Фенотипическая изменчивость проявляется в виде модификаций - это изменение свойств клетки под влиянием внешних воздействий.
Модификации могут быть длительными и кратковременными. Модификационные изменения касаются подавляющего большинства клеток популяции.
Генотипическая - это мутации или рекомбинации. Мутации могут быть спонтанными и индуцированными.
Рекомбинации - это взаимодействие между двумя геномами, обладающими различными генотипами, которое приводит к образованию генома, сочетающего гены донора и реципиента. В процессе рекомбинации бактерий условно делят на клетки-доноры, которые передают генетический материал, и клетки-реципиенты, которые его принимают. Рекомбинация у бактерий рассматриваются как аналоги полового размножения.
Особенности рекомбинаций у бактерий:
- отсутствует мейоз. Образуется не зигота, а меразигота.
- всегда направлена от донора к реципиенту.
- количество генетического материала у рекомбинанта всегда больше одного. Рекомбинант содержит всю генетическую информацию реципиента и часть
генетической информации донора.
У эукариот механизм рекомбинации один – мейоз; у бактерий различают три вида рекомбинаций:
1) трансформацияэто обмен генетической информации с помощью чистой ДНК.
2) трансдукция – этот способ переноса генетической информации с помощью фагов.
3) конъюгация – это способ передачи генетической информации, когда между двумя бактериями образуются цитоплазмические мостики. При конъюгации
в клетку-реципиент может перейти почти весь геном.
Генетические методы применяются в практических целях как для обнаружения микроба в исследуемом материале без выделения чистой культуры, так и для определения таксономического положения микроба и проведения внутривидовой идентификации.
Секвенирование генома – определение последовательности пар нуклеотидов ДНК.
Рестрикционный анализ – этот метод основан на применении ферментов рестриктаз – это эндонуклеазы, которые расщепляют молекулу ДНК только в определённых местах. Если выделенную из конкретного материала ДНК обработать определенной рестриктазой, то это приведет к образованию строго определенного количества фрагментов ДНК фиксированных размеров.
Риботипирование – позволяет определить вид бактерий. Последовательность нуклеотидных оснований в оперонах, кодирующих рРНК, характеризуется наличием как консервативных участков, которые имеют сходное строение у различных бактерий, так и вариабельных последовательностей, которые родо- и видоспецифичны и являются маркерами при генетической идентификации.
Молекулярная гибридизация – применяется в геносистематике. Этот метод позволяет выявить степень сходства различных ДНК.
ПЦР используется для обнаружения генов или соответствующих нуклеотидных последовательностей, кодирующих либо видовую принадлежность, либо иной признак.
Метод ПЦР основан на принципе комплементарности и позволяет увеличивать (амплифицировать) количество исследуемого образца ДНК. Этот метод обладает чрезвычайно высокой чувствительностью и теоретически позволяет обнаружить в исследуемом материале даже единичные молекулы ДНК.
Разновидности ПЦР:
- ПЦР в режиме реального времени; даёт возможность определить количество фрагментов ДНК находящегося в материале, т.е. проводить количественный анализ;
- мультиплексная ПЦР – преимущество заключается в том, что в реакционную смесь можно вводить 2 – 4 и более пары праймеров. Они характерны для различных возбудителей.
- обратнотранскрипционная ПЦР – позволяет осуществить копирование РНК возбудителей.
ДНК – чипы являются новейшими технологиями гибридизационных методов молекулярно-генетического анализа. Они представляют собой носители известных олигонуклеотидов (менее 20 оснований каждый), комлементарных участкам исследуемого генома (или геномов) и занимающих определенный сайт (ячейку). При наличии в исследуемой пробе фрагментов искомой ДНК они гибридизуются (соединяются по принципу комплементарности) с нуклеотидными последовательностями, сидящими на чипе.
Классификация бактерий.
Основной таксономической единицей у бактерий является вид. Для обозначения вида у бактерий используется двойная (бинарная) номенклатура
Вид у бактерийэто совокупность родственных бактерий, которые обладают сходными биологическими свойствами и имеют общее происхождение. В настоящее время существует 3 подхода к классификации бактерий:
1. Рутинная классификация.
Она лежит в основе определителя бактерий под редакцией Берджи.
2. Нумерическая таксономия.
3. Геносистематика.
Заключение: студенты ознакомлены с основами генетики бактерий, с основными методами генодиагностики.
ДНК-содержащие вирусы имеют либо собственные ферменты репликации (в капсиде), либо в их геноме закодирована информация о синтезе вирусных ферментов, обеспечивающих репликацию вирусной нуклеиновой кислоты. Количество этих ферментов различно в применении к разным вирусам. Например, в геноме бактериального вируса Т4 закодирована информация о синтезе около 30 вирусных ферментов. Далее геном крупных вирусов кодирует нуклеазы, разрушающие ДНК клетки-хозяина,а также белки, воздействие которых на клеточную РНК-полимеразу сопровождается тем, что «обработанная таким образом РНК-полимераза транскрибирует на разных стадиях вирусной инфекции разные вирусные гены. Напротив, малые по размерам ДНК-содержащие вирусы в большей мере зависят от ферментов клеток-хозяев. Например, синтез ДНК аденовирусов обеспечивается клеточными ферментами.[ ...]
Бактериальные ДНК - это высокополимерные соединения, состоящие из большого числа нуклеотидов - полинуклеотиды с молекулярным весом около 4 млн. Молекула ДНК представляет собой цепь нуклеотидов, где расположение их имеет определенную последовательность. В последовательности расположения азотистых оснований закодирована генетическая информация каждого вида. Нарушение этой последовательности возможно при естественных мутациях или же под влиянием мутагенных факторов. При этом микроорганизм приобретает или утрачивает какое-либо свойство. У него наследственно изменяются признаки, т. е. появляется новая форма микроорганизма. У всех микроорганизмов - прокариотов и эукариотов - носителями генетической информации являются нуклеиновые кислоты - ДНК и РНК. Лишь некоторые вирусы представляют собой исключение: у них ДНК отсутствует, а наследственная информация записана или отражена только в РНК.[ ...]
В бактериальных клетках в общем количестве оснований ДНК 32-65 мол.% гуанина и цитозина.[ ...]
Ядро бактериальной клетки. Примерно 1-2% веса сухой массы микроорганизмов приходится на ДНК, в которой заложена генетическая информация организма. У большинства микроорганизмов имеются области (или несколько областей), в которой сконцентрировано основное количество ДНК, имеющие определенную структуру (или органеллу) и называющиеся ядром. Ядро (или ядерное вещество) связано с цитоплазматической мембраной, независимо от того, окружено оно элементарными мембранами (как у амебы) или не имеет их (как у бактерий и сине-зеленых водорослей). Ядерное вещество активизируется в период размножения и при наступлении возрастных изменений, связанных со старением клетки.[ ...]
Сегмент ДНК (ген), который предназначен для молекулярного клонирования, должен обладать способностью к репликации при переносе его в бактериальную клетку, т. е. быть репликоном. Однако он такой способностью не обладает. Поэтому, чтобы обеспечить перенос и обнаружение клонируемых генов в клетках, их объединяют с так называемыми генетическими векторами. Последние должны обладать, как минимум, двумя свойствами. Во-первых, векторы должны быть способны к репликации в клетках, причем в нескольких копиях. Во-вторых, они должны обеспечивать возможность селекции клеток, содержащих вектор, т. е. обладать маркером, на который можно вести контрселекцию клеток, содержащих вектор вместе с клонируемым геном (рекомбинантные молекулы ДНК). Таким требованиям отвечают плазмиды и фаги. Плазмиды являются хорошими векторами по той причине, что они являются репликонами и могут содержать гены резистентности к,какому-либо антибиотику, что позволяет вести селекцию бактерий на устойчивость к этому антибиотику и, следовательно, легкое обнаружение рекомбинантных молекул ДНК.[ ...]
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками - гистонами - ив нуклео-иде расположена в виде пучка фибрилл.[ ...]
Применение методов рекомбинантной ДНК для получения биологических агентов для борьбы с загрязнениями находится на ранней стадии, но есть метод, который может оказаться полезным в обозримом будущем - это генетическое зондирование. Отбор организмов, способных трансформировать новое соединение, часто основан на способности использовать вещество как субстрат роста. Если рост слабый или субстрат только комета-болизируется, то методы селекции окажутся непригодными для идентификации деградативной способности. Следовательно, было бы полезно развивать генетическое зондирование для определения специфических последовательностей в плазмидах и хромосомах, это необходимо для определения катаболического потенциала, даже если этот потенциал не экспрессируется. Такие зонды разработаны для TOL-плазмид. Метод может определить одну бактериальную колонию, содержащую TOL-плазмиду, среди 106 колоний Escherichia coli. Такой мощный инструмент будет иметь огромное значение для выделения скрытых катаболических функций .[ ...]
Разработка изящной методики «клонирования» ДНК для получения большого количества точных копий специфических фрагментов ДНК (рис. 13.4) открыла в последнее время новые горизонты в изучении структуры, организации и функции генома. Если расщепить двухцепочечную ДИК одним из ферментов «рестрикции» (одной из нуклеаз), специфично узнающих и расщепляющих короткие последовательности нуклеотидов (4- 6 пар), то возникают в высшей степени воспроизводимые фрагменты ДНК. Концы двух цепей ДНК обычно бывают смещены относительно друг друга вследствие специфичности мест разрезания двухцепочечной молекулы, цепи которой комплементарны по составу оснований. ДНК обычно встраивают в плазмидный ген, важный для селекции, такой, как ген устойчивости к антибиотикам, что позволяет содержащим такую плазмиду бактериям расти в присутствии антибиотика.[ ...]
В бактериях при репликации образуется много копий плазмид, и таким образом можно «вырастить» большие количества встроенных фрагментов ДНК, а затем снова просто выделить их путем расщепления тем же самым ферментом рестрикции с разделением полученных продуктов гель-электрофорезом. Использование этого метода рекомбинации ДНК произвело революцию в изучении генов.[ ...]
Недавно было обнаружено, что мутагенное действие на бактериальные-ДНК-вирусы оказывают лучи с длиной волны 320-400 нм (область, близкая к зоне видимого света), обладающие низкой интенсивностью . Возможное влияние радиации в этом диапазоне длин волн на вирусы растений пока не обнаружено.[ ...]
Кривые зависимости реассоциацни от СОТ, полученные для бактериальной ДНК, лишены перегибов, а ДИК эукариот реас-социирует по другому типу (рис. 13.2). При низких концентрациях ДНК и коротком времени инкубации реиатурирует заметная доля одноцепочечной ДНК, а при увеличении СОТ образуется дополнительное количество двухцепочечных молекул, так что получается двухфазная кривая. Быстрая реиатурация при низких значениях СОТ показывает, что какие-то последовательности у эукариот повторяются много раз, т. е. до 10 000 раз и более.[ ...]
Отсутствие СХС может иммитироваться и в тех случаях, когда ДНК тестерных фагов не содержат сайтов, узнаваемых существующей в исследуемом штамме рестриктазой. Это явление представляет собой один из вариантов эволюционных адаптивных изменений бактериальных вирусов, призванных способствовать преодолению ими барьера СХС. Действие давления отбора в данном конкретном случае выражается в статистическом достоверном уменьшении числа или даже полной элиминации в фаговой ДНК последовательностей нуклеотидов, являющихся субстратом рестриктаз, характерных для клеток-хозяев бактериального вируса .[ ...]
Линдегрен описал возможные стадии образования бактериофага из ДНК профага, предположив, что профаг возникает как фрагмент чужеродной бактериальной ДНК, случайно проникший в клетку, который на ранних стадиях делится синхронно с бактериальной ДНК. Следующим важным этапом в развитии вируса явилось бы такое изменение профага, в результате которого стало возможным его независимое от ДНК клетки-хозяина размножение; в результате профаг использовал бы. все доступные нуклеотиды, нарушив тем-самым рост клетки-хозяина. Наконец, на какой-то более поздней стадии могла бы образоваться защитная белковая оболочка и возникли другие белки, что должно было обеспечить выживание ДНК вне организма хозяина и эффективное заражение новых клеток. Отделившийся фрагмент бактериальной ДНК вначале, очевидно, кодировал белки, приспособленные к бактериальным функциям. Необходимы очень существенные изменения в ДНК, чтобы могли возникнуть объекты настолько сложные и специализированные, как, скажем, фаг Т2 Е. coli, содержащие к тому же основания, которые в бактериальной ДИК отсутствуют.[ ...]
Генетическая информация бактерий не ограничивается ДНК, расположенной в нуклеоиде бактериальной клетки. Как уже отмечалось в предыдущих разделах книги, носителями наследственных свойств служат также внехромосомные элементы, получившие общее название плазмид. В отличие от ДНК ядерных эквивалентов-нуклеоидов, являющихся органоидами бактериальной клетки, плазмиды представляют собой независимые генетические элементы. Потеря плазмид или их приобретение не отражается на биологии клетки (приобретение плазмид оказывает положительное влияние лишь на популяцию в целом, повышая жизнеспособность вида). К трансмиссивным относят плазмиды, инициирующие свойства доноров у клеток-хозяев. При этом последние получают новое качество - возможность конъюгировать с клетками-реципиентами и отдавать им свои плазмиды. Клетки-реципиенты, приобретая во время конъюгации плазмиды, сами превращаются в доноров.[ ...]
Отсутствие адсорбции не исчерпывает разнообразия вариантов взаимодействия бактериальных вирусов и микробных клеток. Они иллюстрируют лишь одну сторону этого явления, а именно проявление клеточных защитных механизмов, фенотипически (по критерию отсутствия роста) иммитирующих рестрикцию. Однако, существует и другой вариант взаимодействия клетка-бактериофаг, который может иммитировать отсутствие СХС. Примерами таких механизмов является синтез ингибиторов и метилаз кодируемых фаговыми генами, защищающих вирусную ДНК от действия рестриктаз II типа.[ ...]
Механизм обеззараживающего действия хлора связан с нарушением обмена веществ бактериальной клетки в процессе дезинфекции воды. При этом выявлено влияние на ферментную активность бактерий, в частности, на дегидрогеназы, катализирующие окислительно-восстановительные реакции в бактериальной клетке. А. М. Ски-дальской (1969) было изучено влияние хлора на процесс декарбоксилирования аминокислот бактерий, протекающий в присутствии строго специфичных ферментов-декарбоксилаз, а также был определен нуклеотидный состав ДНК кишечной палочки после окончания процесса обеззараживания при различных уровнях бактерицидного эффекта.[ ...]
Бактериофаги Т-группы имеют форму барабанных палочек размером 100 х 25 нм. Их геном представлен ДНК. Они являются вирулентными фагами, т. к. после инфицирования ими бактериальных клеток последние лизируются с освобождением большого количества вновь синтезированных фаговых частиц.[ ...]
Плазмиды бактерий - это генетические структуры, находящиеся в цитоплазме и представляющие собой молекулы ДНК размером от 2250 до 400 ООО пар азотистых оснований. Они существуют обособленно от хромосом в количестве от одной до нескольких десятков копий на одну бактериальную клетку.[ ...]
Штамм Pseu.dom.onas вуг1 ае ри. рка8еоИсо1а обладает плазмидой длиной 150 тыс. п. н., которая может реплицироваться автономно, а может интегрироваться в бактериальную хромосому. Последующее неточное вырезание позволило получить семейство плазмид длиной от 35 до 270 тыс. п. н., некоторые из них содержали большие сегменты хромосомной ДНК .[ ...]
В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты (эндонуклеазы), которые стали частью клеточной (бактериальной) системы рестрикции-модификации. У бактерий системы рестрикции-модификации являются внутриклеточной иммунной системой защиты от чужеродной ДНК. В отличие от высших организмов, у которых распознание и разрушение вирусов, бактерий и других патогенов происходит внеклеточно, у бактерий защита от чужеродной ДНК (ДНК растений и животных, в организме которых они обитают) происходит внухриклеточно, т. е. тогда, когда чужеродная ДНК проникает в цитоплазму бактерий. С целью защиты бактерии в ходе эволюции развили также способность «метить» собственную ДНК метилирующими основаниями на определенных последовательностях. По этой причине чужеродная ДНК из-за отсутствия в ней метальных групп на тех же последовательностях плавится (разрезается) на фрагменты разными бактериальными рестриктазами, а затем деградируется бактериальными экзонуклеазами до нуклеотидов. Можно сказать, что таким образом бактерии защищают себя от ДНК растений и животных, в организме которых они обитают временно (как патогены) или постоянно (как сапрофиты).[ ...]
Наследственные свойства бактерий или отдельные признаки закодированы в единицах наследственности - генах, линейно расположенных в хромосоме вдоль нити ДНК. Следовательно, ген является фрагментом нити ДНК- Каждому признаку соответствует определенный ген, а часто еще меньший отрезок ДНК - кодон. Иначе говоря, в нити ДНК в линейном порядке расположена информация обо всех свойствах бактерий. При этом у бактерий есть еще одна особенность. В ядрах эукариотов содержится обычно несколько хромосом, число их в ядре постоянно у каждого вида. Нуклеоид бактерий содержит лишь одно кольцо из нити ДНК, т. е. одну хромосому. Однако запасом информации, заключенным в одной хромосоме или в кольцеобразно сомкнувшейся двунитчатой спирали ДНК, сумма наследственных признаков бактериальной клетки не исчерпывается. Плазмиды содержат ДНК, также несущую генетическую информацию, передаваемую от материнской клетки к дочерней.[ ...]
Мутации - это изменения в генном аппарате клетки, которые сопровождаются изменениями контролируемых этими генами признаков. Различают макро- и микроповреждения ДНК, ведущие к изменению свойств клетки. Макроизменения, а именно: выпадение участка ДНК (деления), перемещение отдельного участка (транслокация) или поворот определенного участка молекулы на 180° (инверсия) -у бактерий наблюдаются сравнительно редко Гораздо более характерны для них микроповреждения, или точечные мутации, т. е. качественные изменения в отдельных генах, например замена пары азотистых оснований. Мутации бывают прямые и обратные, или реверсивные. Прямые - это мутации организмов дикого типа, например утрата способности самостоятельно синтезировать факторы роста, т. е. переход от прото- к ауксотрофности. Обратные мутации представляют собой возвращение, или реверсию, к дикому типу. Способность к реверсии характерна для точечных мутаций. В результате мутаций изменяются такие важнейшие признаки, как способность самостоятельно синтезировать аминокислоты и витамины (ауксотрофные мутанты), способность к образованию ферментов. Эти мутации называют биохимическими. Хорошо известны также мутации, ведущие к изменению чувствительности к антибиотикам и другим антимикробным веществам. По происхождению мутации разделяют на спонтанные и индуцированные. Спонтанные возникают самопроизвольно без вмешательства человека и носят случайный характер. Частота таких мутаций очень низка и составляет от 1 X Ю“4 ло 1 X 10-10. Индуцированные возникают при воздействии на микроорганизмы физических или химических мутагенных факторов. К физическим факторам, обладающим мутагенным действием, относятся ультрафиолетовое и ионизирующие излучения, а также температура. Химическими мутагенами являются ряд соединений и среди них наиболее активны так называемые супермутагены. В природных условиях и эксперименте изменения в составе бактериальных популяций могут возникать в результате действия двух факторов - мутаций и автоселекции, происходящей в результате адаптации некоторых мутантов к условиям среды обитания. Такой процесс, очевидно, наблюдается в среде, где преобладающим источником питания является синтетическое вещество, например, ПАВ или капролактам.[ ...]
Одиночная клетка E. coli окружена трехслойной клеточной оболочкой толщиной порядка 40 нм, представляющей собой «мешок» или «конверт», в котором заключено клеточное содержимое в виде, примерно, 2 х Ю 1Я г белка, 6 х 10 16 г ДНК и 2 х 10 14 г РНК (в основном ри-босомной РНК). В бактериальной клетке синтезируется около 2000 разных белков, большинство которых содержится в цитоплазме. Концентрация одних белков составляет 10“® М, тогда как других - порядка 2 х 10"4 М (от 10 до 200 000 молекул на клетку).[ ...]
У одноклеточных организмов половое размножение существует в нескольких формах. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.[ ...]
Бактерии: прокариоты («доядерные») одноклеточные организмы. Их клетки не имеют отделенного от цитоплазмы ядра. Однако генетическая программа, как и у всех живых организмов, закодирована в виде последовательности нуклеотидов в ДНК и несет информацию о структуре белков. Бактериальные клетки не содержат таких органелл, как хлоропласты (специализированные для фотосинтеза) и митохондрии (специализированные для клеточного дыхания и синтеза АТФ). Эти биохимические процессы происходят у бактерий в цитоплазме.[ ...]
Крайне малые размеры клеток являются характерной, но не главной особенностью бактерий. Все бактерии представлены особым типом клеток, лишенных истинного ядра, окруженного ядерной мембраной. Аналогом ядра у бактерий является нуклеоид - ДНК-содержащая плазма, не отграниченная от цитоплазмы мембраной. Кроме того, для бактериальных клеток характерны отсутствие митохондрий, хлоропла-стов, а также особое строение и состав мембранных структур и клеточных стенок. Организмы, в клетках которых отсутствует истинное ядро, называются прокариотами (доядер-ными) или протоцитами (т. е. организмами с примитивной организацией клеток).[ ...]
Микоплазменные клетки имеют овальную форму, а их размеры составляют около 0,1-0,25 нм в диаметре (рис. 43). Для них характерно наличие тонкой наружной плазматической мембраны (толщина - около 8 нм), которая окружает цитоплазму, содержащую молекулу ДНК, достаточную для кодирования около 800 разных белков, РНК разных типов, рибосом диаметром порядка 20 нм. В их цитоплазме содержатся различные включения в виде белков, гранул липидов и других соединений. Из-за недостаточной жесткости клеточкой мембраны микоплазмы проходят через бактериальные фильтры.[ ...]
Установлено, что на рибосомах происходят связывание активированных аминокислот и укладка их в полипептидную цепь в соответствии с генетической информацией, полученной из ядра через информационную (матричную) РНК (мРНК), которая как бы считывает соответствующую информацию с ДНК и передает ее на рибосомы. Целый ряд белков синтезирован на изолированных рибосомах и при этом отмечено включение в них меченых аминокислот. Роль матрицы в белковом синтезе выполняет мРНК, которая прикрепляется к рибосоме. На поверхности последней происходит взаимодействие между комплексом аминокислот, транспортной РНК, несущей очередную аминокислоту, и нуклеотидной последовательностью информационной РНК, которая функционирует на рибосоме однократно и после синтеза полипептидной цепи распадается, а вновь синтезированный белок накапливается в рибосомах. В бактериальной клетке при периоде регенерации 90 мин скорость кругооборота мРНК достигает 4-6 с.[ ...]
Цитоплазма представляет собой коллоидный раствор, дисперсной фазой которого являются сложные белковые соединения и вещества, близкие к жирам, а дисперсионной средой - вода. У некоторых форм бактерий в цитоплазме содержатся включения - капельки жира, серы, гликогена и др. Постоянными составляющими бактериальных клеток являются особые выросты цитоплазматической мембраны - мезосомы, в которых содержатся ферментные окислительно-восстановительные системы. В этих образованиях идут в основном процессы, связанные с дыханием бактерий. В мелких включениях - рибосомах, содержащих рибонуклеиновую кислоту, осуществляется биосинтез белка. Большинство видов бактерий не имеет обособленного ядра. Ядерное вещество, представленное ДНК, у них не отделено от цитоплазмы и образует нуклеоид. Транспортировка веществ, необходимых для жизнедеятельности клетки, и отвод продуктов обмена осуществляется по особым каналам и полостям, отделенным от цитоплазмы мембраной, имеющей такое же строение, как и цитоплазматическая. Это структурное образование называется эндоплазматической сетью (ретикулум).[ ...]
Представление об изменчивости и наследственности бактерий нельзя составить без знания некоторых положений молекулярной генетики прокариотической клетки. В основе процессов приспособления микробных культур к изменяющимся экологическим условиям лежат изменчивость и наследственность, являющиеся разделами генетики бактерий. При изложении цитологии бактериальной клетки уже рассматривалась структура ДНК и РНК и их роль в жизни клетки. Характерное строение ДНК сохраняется у каждого вида и передается потомству из поколения в поколение, как и другие признаки. ДНК бактерий представляет собой двунитчатую спираль, замыкающуюся в кольцо. Кольчатая нить ДНК бактерий, расположенная в ну-клеоиде, не содержит белка. Такое кольцо ДНК соответствует хромосоме эукариотической клетки. Известно, что в хромосоме эукариотических клеток, кроме ДНК, всегда содержится белковый компонент. Отсюда следует, что понятие хромосомы у эукариотов несколько отлично от понятия хромосомы бактерий. Нить ДНК, представляющая собой хромосому бактерий, разумеется, у разных видов различается. Сахарофосфатный компонент ДНК У всех видов бактерий одинаков; расположение азотистых оснований и их комбинация, напротив, различаются у разных видов.[ ...]
Все возрастающее беспорядочное применение антибиотиков в животноводстве, которые используются в малых дозах как стимуляторы роста, а также в качестве превентивной меры против вызванных стрессом желудочно-кишечных расстройств у животных на фермах, приводит к все более широкому распространению в микробных популяциях R-фактора устойчивости к антибиотикам, передающегося от одной бактериальной клетки к другой при конъюгации. Передача происходит через плазмиду, которая представляет собой кольцевую экстрахромосомную ДНК, способную к репликации.[ ...]
В противоположность вирулентным фагам, известны так называемые фаги умеренного действия, или просто умеренные фаги. Типичным представителем таких фагов является фаг X, который тоже использовался и используется в качестве экспериментальной модели для выяснения многих вопросов молекулярной генетики. Фагу X присущи два важных свойства. Подобно вирулентным фагам он может инфицировать бактериальные клетки, размножаться вегетативно, продуцируя в клетках сотни копий и лизировать клетки С освобождением зрелых фагочастиц. Однако ДНК этого фага может включаться в бактериальную хромосому, превращаясь в профаг. При этом происходит так называемая лизогенизация бактерий, а бактерии, содержащие профаг, называют лизогенными. Лизогенные бактериальные клетки могут обладать профагом бесконечно долгое время, при этом не лизируясь. Лизис с освобождением новых фагочастиц отмечается после воздействия на лизогенные бактерии какого-либо фактора, например УФ-излучения, которое индуцирует развитие профага в фаг. Изучение лизогенных бактерий позволило получить ряд новых данных о роли разных белков в действии фаговых генов.[ ...]
Геном хлоропластов ряда высших растений состоит из 120 генов. Хлоропластный геном очень сходен с бактериальным геномом как по организации, так и по функциям. В митохондриальном геноме человека, вероятно, отсутствуют интроны, но в ДНК хлоропластов некоторых высших растений, а также в ДНК митохондрий грибов интроны обнаружены. Считают, что хлоропластные геномы высших растений остаются без изменений примерно несколько миллионов лет. Возможно, что такая древность характерна и для митохондриальных геномов млекопитающих, включая человека.[ ...]
Современные схемы, иллюстрирующие работу генов, построены на основании логического анализа экспериментальных данных, полученных с помощью биохимических и генетических методов. Применение тонких электронно-микроскопических методов позволяет в буквальном смысле слова увидеть работу наследственного аппарата клетки. В последнее время получены электронно-микроскопические снимки, на которых видно, как на матрице бактериальной ДНК, в тех участках, где к ДНК прикреплены молекулы РНК-полимеразы (фермента, катализирующего транскрипцию ДНК в РНК), происходит синтез молекул и-РНК. Нити и-РНК, расположенные перпендикулярно к линейной молекуле ДНК, продвигаются вдоль матрицы и увеличиваются в длине. По мере удлинения нитей РНК к ним присоединяются рибосомы, которые, продвигаясь, в свою очередь, вдоль нити РНК по направлению к ДНК, ведут синтез белка.[ ...]
Трансдукция - это перенос генетического материала от бактерии-донора к бактерии-реципиенту с помощью фага. Впервые явление трансдукции было открыто в 1951 г. Ледербергом с сотрудниками у Salmonella typhimurium. Сейчас различают неспецифическую и специфическую трансдукции. При неспецифической трансдукции возможен перенос фагом любого признака от бактерии-донора к бактерии-реципиенту. Перенос осуществляется только умеренными (невирулентными) фагами. Умеренные фаги способны заражать бактерии, однако не размножаются в них и не вызывают лизиса, а включаются в ДНК бактериальной клетки и в таком неинфекционном состоянии в виде так называемого профага передаются от клетки к клетке при размножении. Культуры бактерий, содержащие профаг, называются лизогенными. В этих культурах с небольшой частотой (в одной из 102 - 105 клеток) наблюдается спонтанное размножение фага и происходит лизис клетки с освобождением фаговых частиц, обнаруживаемых с помощью бактерий-индикаторов, для которых такой фаг вирулентен.[ ...]
Опыты проводили на трехкамерной ячейке, состоящей из центральной рабочей и двух электродных камер. В рабочую камеру размером 25 X 7 X 37 мм (длина X ширина X высота), отделенную от электродных целлофановыми мембранами, помещали 750 мг ваты. Через нее снизу вверх подавали с постоянной скоростью исходный раствор исследуемых веществ. За содержанием соединений в исходных, подаваемых в рабочую камеру растворах (С0), и в растворах, выходящих из камеры (Ci), следили по максимумам поглощения белков и нуклеиновых кислот в диапазоне волновых чисел (35,5-38) X Ю3 см-1 с помощью УФ-спектрофотометра Specord UV-VIS. Электродные камеры заполняли гранулированным активированным углем и через них отдельным протоком пропускали дистиллированную воду.
МОРФОЛОГИЯ БАКТЕРИЙ
Бактерии – микроскопические, обычно одноклеточные организмы растительной природы (микрофлора); отдельным видам бактерий с достаточным постоянством присуща определенная морфология. Выделяют три основные формы бактерий – шаровидные или овальные (кокки), палочки (бациллы) и спиралевидные.
Кокки подразделяют на парные – диплококки (нейссерии); тетракокки, располагающиеся по 4 в форме квадратов; пакетообразующие кокки, или сарцины , располагающиеся «этажами»; стрептококки , располагающиеся цепочками; стафилококки , образующие бесформенные скопления, несколько напоминающие гроздья винограда.
Палочки . Среди палочек выделяют одиночные , беспорядочно расположенные бактерии (энтеробактерии), диплобациллы, располагающиеся попарно (по одной линии), и стрептобациллы, образующие цепочки (палочки сибирской язвы).
Спиралевидные бактерии разделяют на две группы – вибрионы и сходные по форме бактерии, изогнутость тела которых не превышает четверти оборота спирали (кампилобактеры), и спирохеты и спириллы, имеющие изгибы, равные одному или нескольким оборотам спирали (возбудитель сифилиса).
Любая бактерия составлена из трех компонентов: поверхностные структуры, клеточная оболочка, цитоплазма.
Поверхностные структуры бактерий – капсулы, жгутики и микроворсинки.
Капсулы окружают клеточную оболочку многих бактерий, в том числе патогенных. У капсул отсутствует упорядоченная организация, характерная для клеточной оболочки бактерий. Выделяют микрокапсулы, которые выявляют только при электронной микроскопии в виде слоя мукополисахаридных микрофибрилл) и макрокапсулы (обнаруживают при световой микроскопии).
Большинство бактериальных капсул состоит из сложных полисахаридов. Выявляются пои окраске по Бурри-Гинсу, либо с помощью реакции набухания по Нойфельду. Капсулы могут включать азотсодержащие соединения, например у пневмококков (состоят из полисахаридов, глюкозамина и глюкуроновой кислоты), но могут и не содержать азот, например капсулы лейконостоков (состоят из декстрина, левулана, фруктозана и других полимеризованных моносахаров).
Капсулы некоторых бактерий (Bacillus anthracis) состоят из полисахаридов и полипептидов, образованных мономерами D-глутаминовой кислоты, которая защищает бактерию от протеолитических ферментов фагоцитов.
Жгутики присутствуют у многих бактерий и обеспечивают подвижность. Жгутик – спирально изогнутая нить, приводимая во вращение «мотором», находящимся в месте ее прикрепления к мембране. У разных бактерий толщина жгутиков варьирует от 12 до 18 нм, длина может достигать 20 мкм.
Жгутики бактерий состоят из белка (флагеллина) и построены из его субъединиц с относительно малой молекулярной массой. Нити жгутиков приводит в движение мембранный шарнироподобный базальный крюк, закрепленный с помощью базального тельца, состоящего у грамположительных бактерий из одной, а у грамотрицательных бактерий из двух пар колец. Кольца выполняют роль «приводного диска» и «подшипника» на внутренней поверхности пептидогликанового слоя. Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор).
Расположение.
Перитрихи. Жгутики расположены по всей поверхности клеточной стенки (бактерии семейств Enterobacteriaceae и Bacillaceae).
Монотрихи. Один толстый жгутик на одном конце (вибрионы).
Политрихи. Пучок из 2-50 жгутиков, видимый как одиночный.
Полярные жгутики прикреплены к одному или обеим концам бактерии. Лофотрихи – пучок жгутиков на одном конце бактерии (Pseudomonas). Амфитрихи – биполярно расположенные пучки (Spirillum).
Микроворсинки (пили, фимбрии) это белковые волоски (от 10 до нескольких тысяч) толщиной 3-25 нм и длиной до 12 мкм.
А. Обыкновенные пили. Многие грамотрицательные бактерии имеют длинные и тонкие пили (фимбрии), начинающиеся на цитоплазматической мембране и пронизывающие клеточную стенку. Они образованы белками одного типа, молекулы которых формируют спиральную нить. Их основная функция – прикрепление бактерий к субстратам , например поверхности слизистых оболочек, что является важным фактором колонизации и инфицирования. Кроме того, увеличение площади поверхности бактериальной клетки дает ей дополнительные преимущества в утилизации питательных веществ окружающей среды.
Б. F- пили (фактор фертильности) – специальные образования, участвующие в коньюгации бактерий. Имеют вид полых белковых трубочек длиной 0,5-10 мкм. Их образование кодируется плазмидами.
Клеточная оболочка большинства бактерий состоит из клеточной стенки и находящейся под ней цитоплазматической мембраны.
Клеточная стенка бактерий тонкая, эластичная и ригидная, может полностью отсутствовать у некоторых бактерий (например, L-форм и микоплазм). Клеточная стенка защищает бактерии от внешних воздействий, придает им характерную форму, через нее осуществляется транспорт питательных веществ и выделение метаболитов. На ее поверхности располагаются разнообразные рецепторы для бактериофагов, бактериоцинов и различных химических веществ. КС поддерживает постоянство внутренней среды и выдерживает значительное давление изнутри (например, парциальное давление внутриклеточных веществ грамположительных бактерий может достигать 30 атмосфер). Структура и состав элементов КС определяют способность воспринимать красители, т.е. их тинкториальные свойства . В основу одного из основных принципов дифференциации бактерий положена способность воспринимать и удерживать внутри клетки красящий комплекс генцианового фиолетового с йодом, либо терять его после обработки спиртом (окраска по Граму). Соответственно выделяют грамположительные (окрашиваются в фиолетово0пурпурный цвет) и грамотрицательные (красного цвета).
Основной компонент КС бактерий – пептидогликан (муреин). Пептидогликана относительно больше в грамположительных бактериях: доля муреиновой сети толщиной примерно в 40 слоев составляет 30-70% сухой массы КС. Грамотрицательные бактерии содержат всего 1-2 слоя муреина, составляющего около 10% сухой массы КС.
Пептидогликан представлен полимерными молекулами, состоящими из повторяющихся дисахаридных групп, в образовании которых участвуют N -ацетилглюкозамин и N -ацетилмурамовая кислота , последняя связывает дисахариды с олигопептидами (из 20 известных аминокислот в КС бактерий найдены лишь 4 – глутаминовая кислота, глицин, лизин и аланин). В состав КС бактерий также входят уникальные аминокислоты, например диаминопимелиновая и D-изомеры глутаминовой кислоты и аланина. Лизоцим гидролизует пептидогликан, расщепляя гликозидные связи между N-ацетилглюкозамином и N-ацетилмурамовой кислотой.
Перекрестное связывание пептидогликана заключается в образовании пептидной связи между терминальным остатком боковой пептидной цепи (обычно D-аланином) с предпоследним остатком примыкающей боковой цепи (L-лизином или диаминопимелиновой кислотой).
Грамположительные бактерии имеют несложно организованную, но мощную КС, состоящую преимущественно из множественных слоев пептидогликана, включающих уникальные полимеры тейхоевых кислот – цепи из 8-50 остатков глицерина или рибита, связанные между собой фосфатными мостиками.
Грамотрицательные бактерии имеют более тонкую (по сравнению с грамположительными бактериями) КС, включающую бимолекулярный слой пептидогликана и не содержащую тейхоевой кислоты.
Поверх пептидогликанового слоя расположена дополнительная, или внешняя мембрана. Её толщина превышает размеры монослоя пептидогликана.
Компоненты внешней мембраны: фосфолипидный бислой, белки, полисахариды и ЛПС, расположенные мозаично.
Фосфолипидный бислой прикреплен к пептидогликану липопротеинами, пересекающими периплазматическое пространство.
Белки , в том числе порины , образующие трансмембранные каналы, вовлечены в транспорт ионов и гидрофильных соединений из внешней среды в периплазму.
ЛПС образован из липидной части (липид А), насыщенной полисахаридами сердцевины и боковых полисахаридных цепей. Полисахаридная часть ЛПС обладает иммуногенными свойствами и называется О-Аг. Липидная часть термоустойчива и отвечает за биологические эффекты эндотоксина.
Аутолизины . КС бактерий содержат аутолизины – ферменты, растворяющие пептидогликановый слой. Их активность необходима для процессов роста КС, разделения клеток, споруляции и достижения состояния компетентности при трансформации.
Цитоплазматическая мембрана (иначе клеточная, или плазматическая мембрана) – физический, осмотический и метаболический барьер между внутренним содержимым бактериальной клетки и внешней средой. ЦПМ имеет сложную трехслойную структуру, для неё характерна выраженная избирательная проницаемость. У некоторых бактерий между ЦПМ и КС располагается периплазматическое пространство – полость, заполненная ферментами (рибонуклеазы, фосфатазы, пенициллиназы и др.), у грамотрицательных бактерий ферменты свободно изливаются в окружающую среду. ЦПМ бактерий состоит из белков, липидов, углеводов и РНК.
Белки ЦПМ разделяют на структурные и функциональные. Последние включают ферменты, участвующие в синтетических реакциях на поверхности мембраны, окислительно-восстановительных процессах, а также некоторые специальные энзимы (например, пермеазы ).
В ЦПМ расположена система электронного транспорта бактерий, обеспечивающая энергетические потребности.
Мезосомы – сложные инвагинации ЦПМ, функции которых до сих пор полностью не установлены. Известно, что они ассоциированы с нуклеоидом и имеют отношение к делению клеток и спорообразованию.
Удаление КС, защищающей прилежащую ЦПМ, приводит к лизису бактерий либо к образованию протопластов и сферопластов, различающихся по происхождению (из грамположительных или грамотрицательных бактерий соответственно), а также по осмотической устойчивости. Пребывая в изотонической среде, бактерии, лишенные КС, способны поглощать О 2 и выделять СО 2 , а также размножаться.
L -формы. Под влиянием некоторых внешних факторов бактерии способны терять КС, образуя L-формы (названы в честь Института им. Д.Листера, где были впервые выделены). Подобная трансформация может быть спонтанной (например, у хламидий) или индуцированной (например, под действием антибиотиков). Выделяют стабильные и нестабильные L -формы. Первые не способны к реверсии, а вторые реверсируют в исходные формы после удаления причинного фактора.
Представители группы микоплазм (класс Mollicutes) не имеют клеточных стенок.
Цитоплазма бактерий – матрикс для реализации жизненно важных реакций – отделена от КС цитоплпзматической мембраной. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза, её основные составляющие – растворимые ферменты и РНК (матричные и транспортные РНК). Разнообразные органеллы, характерные для эукариотических клеток, у бактерий отсутствуют, а их функции выполняет бактериальная ЦПМ.
ДНК . В бактериальной клетке нет ядерной мембраны. ДНК сконцентрирована в цитоплазме в виде клубка, называемого нуклеоидом, или генофором.
Генофор бактерий представлен двойной спиральной кольцевой ковалентно замкнутой суперспирализованной молекулой ДНК, составляющей 2-3% сухой массы клетки (более 10% по объему). Длина контура молекулы варьирует от 0,25 до 3 мм. Суперспираль бактериальной ДНК не содержит гистонов. Объем генетической информации, кодируемой в генофоре, различается между видами (например, геном Escherichia coli кодирует примерно 4 000 различных полипептидов).
Плазмиды . У бактерий может присутствовать дополнительная молекула ДНК в виде внехромосомных элементов либо интегрированных в генофор. Подобные включения называют плазмидами (соответственно эписомальные или интегрированные ). Для ДНК эписом тоже характерна кольцевая форма, но по размеру эписомы меньше бактериальной хромосомы. Плазмиды несут ряд различных генов и часто определяют вирулентность бактерий, но информация, содержащаяся в плазмидах, не является абсолютно необходимой для бактериальной клетки.
Рибосомы бактерий – сложные глобулярные образования, состоящие из различных молекул РНК и многих связанных с ними белков. Всё образование функционирует как локус синтеза белков.
70 S рибосомы . Диаметр бактериальных рибосом около 20 нм. Коэффициент седиментации – 70S (единиц Сведберга). Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50S для одной и 30S для другой. Объединение субъединиц происходит перед началом синтеза белка. В зависимости от интенсивности роста бактериальная клетка может содержать от 5 000 до 50 000 рибосом.
Бактериостатические антибиотики (стрептомицин, тетрациклин, левомицетин) ингибируют синтез белка, блокируя некоторые метаболические процессы, протекающие в рибосомах бактерий.
Запасные гранулы содержат временный избыток метаболитов. Наличие и количество гранул изменяются в зависимости от вида бактерий и их метаболической активности. В виде гранул могут запасаться полисахариды (крахмал, гликоген, гранулёза), жиры (триглицериды, сходные с жирами высших животных, запасаются у дрожжей рода Candida; воска – у микобактерий и нокардий; полимеры β-оксимасляной кислоты – например в клетках Bacillus megaterium), полифосфаты (например, волютин, впервые обнаруженный у Spirillum volutans), сера (у бактерий, окисляющих сульфид до сульфата), белки – например, протоксин (у Bacillus thuringiensis и родственных видов).
Основной секрет органической жизни кроется в способности к размножению и передаче наследственной информации от предыдущих поколений потомкам через довольно простой механизм самокопирования макромолекулы ДНК каждой живой клетки. Каждой, независимо от того, состоит организм из большого количества клеток или же речь идет о тех ДНК, которые находятся в клетках бактерий, этих одноклеточных простейших организмов, не всегда способных даже в большую колонию собраться.
Как у всех представителей органической жизни, наследственная (генетическая) информация бактерий хранится в их ДНК. Что такое генетическая информация? Какая структура хранит наследственную информацию?
- Генетическая информация – это определенная последовательность нуклеотидов. Другого секрета в ядре нет. Копируя эту последовательность, клетка синтезирует самые разнообразные белки. Они же решают все остальные вопросы организма, начиная с организационных, заканчивая снабжением клетки строительным материалом.
- Макромолекула ДНК – четыре нуклеиновых основания (аденин, гуанин, тимин и цитозин), объединенные в двойную спираль сахаром дезоксирибозой и остатками фосфорной кислоты. Именно нуклеиновые основания кодируют последовательность сборки белков независимо от того, есть оформленное ядро в клетке или нет.
Дезоксирибонуклеиновая кислота бактерий имеет такое же строение, как молекулы – хранители наследственной информации всех остальных живых существ на планете. Так же, как все другие органические клетки, бактерия образует из ДНК хромосомы. Но это не значит, что других отличий нет.
Фундаментальным отличием бактерии является то, что у нее нет клеточного ядра, наследственная информация бактерии не собрана в клеточное ядро, это просто кольцевая молекула, которая прилеплена к одной из стенок цитоплазматической мембраны.
 Однако то обстоятельство, что ядра нет, не препятствует активным процессам репликации и трансляции с использованием этого хранителя наследственной информации. Чтобы понять, как происходит передача информации, нужно понимать, что такое хромосомы, гены и клеточное ядро.
Однако то обстоятельство, что ядра нет, не препятствует активным процессам репликации и трансляции с использованием этого хранителя наследственной информации. Чтобы понять, как происходит передача информации, нужно понимать, что такое хромосомы, гены и клеточное ядро.
- Ген – участок макромолекулы, на котором записана последовательность нуклеотидов, позволяющая собирать один определенный вид белка. Другой информации в генах нет.
- Хромосома – комбинация цепи ДНК с белками гистонами, которые ее структурируют и придают ей определенную форму перед тем, как клетка начинает делиться. В фазе, когда деление не происходит, в клетке (или в ядре, если речь идет о ядерных эукариотах) как таковых хромосом нет.
- Клеточное ядро – это клеточная структура, которая содержит наследственную информацию, структурированную в хромосому, когда клетка готовится к делению. В ней инициируется сам процесс деления. Важно помнить, что у бактерий клеточного ядра нет.
Если в эукариотической клетке при делении используются обособленные, специально формирующиеся для удобства деления структуры, то как же происходит размножение бактерий в условиях неоформленного кажущегося сумбура в отсутствие клеточного ядра?
Дезоксирибонуклеиновая кислота бактериальной клетки
Бактериальная молекула ДНК хоть изображается как кольцевая довольно объемная структура, которая располагается в центре клетки, на самом деле представляет собой довольно компактное образование, локализованное на ограниченных участках цитоплазмы.
 Ввиду отсутствия ядерной мембраны, которая бы отгораживала скомпонованную бактериальную макромолекулу от других клеточных структур, генетический аппарат безъядерных организмов нельзя ассоциировать с генетическим аппаратом эукариотов, поэтому генетический аппарат прокариотов назвали нуклеоид.
Ввиду отсутствия ядерной мембраны, которая бы отгораживала скомпонованную бактериальную макромолекулу от других клеточных структур, генетический аппарат безъядерных организмов нельзя ассоциировать с генетическим аппаратом эукариотов, поэтому генетический аппарат прокариотов назвали нуклеоид.
Характерные черты нуклеоида:
- ДНК, в которой содержится нескольких тысяч генов.
- Гены расположены линейно и называются хромосомой. Хромосома бактерии – это линейная совокупность ее генов.
- Макромолекула также сворачивается белками, похожими на эукариотические гистоны.
Нуклеоид крепится к цитоплазматической мембране в тех точках, где начинается и заканчивается репликация (самокопирование).

Экспериментальным путем установлено, что нуклеоид и хромосома – это не одно и то же. Увеличение количества хромосом (линейных генов) – свидетельство того, что бактерии активно делятся. Один нуклеоид может состоять из одной хромосомы или нескольких ее копий. Так, в период деления азотобактерия реплицируется до 20-25 хромосом (копий нуклеоида).
Процесс копирования
В теоретических конструкциях, разработанных микробиологами в те годы, когда изучать сложные молекулярные процессы экспериментальным путем было очень сложно или практически невозможно, копирование дезоксирибонуклеиновой кислоты может осуществляться тремя способами:
- Консервативный, при котором двойная родительская спираль не раскручивается, а двойная дочерняя спираль полностью образовывается из нового материала.
- Дисперсивный, при котором родительская макромолекула распадается на фрагменты, а дочерние формируются на нуклеотидных последовательностях этих фрагментов как на матрицах.
- Полуконсервативный. Согласно этой модели, двойная спираль раскручивается, и каждая цепь спирали служит матрицей для дочерних ДНК. Формируется так называемый гибрид старой макромолекулы и цепи, созданной из новых компонентов.
Когда в 1957 году был найден способ отслеживания процессов, происходящих в бактериальной ДНК при ее репликации, было установлено, что дезоксирибонуклеиновая кислота реплицируется полуконсервативным путем, то есть через раскручивание и использование раскрученных участков в качестве матриц для синтеза новых макромолекул.

Сам процесс репликации бактериальной ДНК очень схож с репликацией ДНК остальных органических механизмов. Происходит он по следующей схеме:
- ДНК-хеликазы раскручивают и разрывают двойную спираль, двигаясь вдоль сахарофосфатного остова дезоксирибонуклеиновой кислоты.
- Ферменты полимеразы катализируют реакции присоединения к однонитевым фрагментам дезоксирибонуклеиновой кислоты комплиментарных нуклеиновых оснований.
После репликации происходит удвоение всех основных частей клетки: органелл, цитоплазматической мембраны, клеточной стенки, и бактериальная клетка распадается надвое.
Проблематика
Помимо исключительно научного интереса в изучении ДНК бактерий, механизм репликации и передачи наследственной информации от одной клетки к другой также имеют исключительную практическую важность.
Широко известный факт, что бактерии очень быстро адаптируются при воздействии на них антибиотиков и начинают выработку определенных белков-антител, которые блокируют разрушительное действие антибиотических средств на клетку бактерии. В следующих поколениях бактерий эта устойчивость к конкретной группе антибактериальных препаратов сохраняется.
 Более того, благодаря горизонтальному переносу генов (не в процессе деления, а в процессе простого контакта одной бактерии с другой) такая генетическая информация также передается, делая устойчивыми к антибиотикам все большее количество видов бактерий.
Более того, благодаря горизонтальному переносу генов (не в процессе деления, а в процессе простого контакта одной бактерии с другой) такая генетическая информация также передается, делая устойчивыми к антибиотикам все большее количество видов бактерий.
Изучением этих свойств бактерий, определением того, как посторонний ген включается в общую структуру дезоксирибонуклеиновой кислоты, и занимается современная микробиология.


